

Downloads
Download



This work is licensed under a Creative Commons Attribution 4.0 International License.
Review
From Bench to Bedside: Current Developments in RNA-Based Therapies for Treatment of Hyperlipidemia
Yufei Zhou and Chen Chen *
Division of Cardiology and Hubei Key Laboratory of Genetics and Molecular Mechanisms of Cardiological Disorders, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China.
* Correspondence: chenchen@tjh.tjmu.edu.cn; Tel. & Fax: 86-27-6937-8422.
Received: 7 October 2022
Accepted: 6 November 2022
Published: 21 December 2022
Abstract: Hyperlipidemia is one of the conditions that constitute metabolic disorder and it is a common public health problem. The condition is characterized by increased levels of cholesterol, triglycerides and/or lipoproteins; it is a recognized as a risk factor for the onset of many diseases such as type 2 diabetes, non-alcoholic fatty liver disease, and cardiovascular disease. Up to now, the primary drugs for treating hyperlipidemia are statins and monoclonal antibody drugs against proprotein convertase subtilisin/kexin type 9 (PCSK9). The main limitation of statins for long-term use is intolerable side effects. Evolocumab and Alirocumab, two monoclonal antibodies against PCSK9, can effectively decrease the level of low-density lipoprotein cholesterol (LDL-C) in patients with statin intolerance and familial hypercholesterolemia, while causing fewer side effects. However, due to its short half-life and high costs, these monoclonal antibody treatments might result in patients’ non-compliance with medication and considerable economic burden on patients. Given that RNA plays a key role in gene regulation, RNA-based therapeutics have become powerful blueprints for designing new anti-hyperlipidemia drugs. Here, we summarized RNA-based therapeutic strategies and the current clinical trials for RNA drugs in hyperlipidemia treatment.
Keywords:
hyperlipidemia microRNA therapeutic strategies clinical trial1. Introduction
Hyperlipidemia has become a common health burden over recent decades, as patients have increasing levels of cholesterol, triglycerides and/or lipoproteins. It is recognized as a risk factor for type 2 diabetes, pancreatitis, non-alcoholic fatty liver disease, chronic kidney disease and cardiovascular diseases [1]. Previous studies showed that lipid metabolism consists of de novo lipogenesis, as well as uptake, oxidation and degradation of lipids, such as fatty acid, triglycerides and cholesterol. Their transportation and degradation are greatly affected by the lipoprotein metabolism, including low-density lipoprotein (LDL), very-low-density lipoprotein (VLDL) and high-density lipoprotein (HDL).
The traditional drugs against hyperlipidemia are mainly small molecule drugs, especially statins and monoclonal antibody drugs. Statins are the first-line lipid-lowering drugs in clinical practice nowadays; they lower the level of LDL-C by inhibiting 3-hydroxy-3-methyl-glutaryl coenzyme-A reductase, a rate-limiting enzyme in cholesterol synthesis. However, with long-term use, some patients suffer from adverse effects; the main reasons for statin intolerance are liver injury [2], statin-associated muscle symptoms (SAMS) [3] and new-onset type 2 diabetes (NOD) [4,5]. Previous studies identified that proprotein convertase subtilisin/kexin type 9 (PCSK9), a serine protease synthesized primarily in hepatocytes, was of great importance to the pathogenesis of familial hypercholesterolemia. PCSK9 acts by inducing the degradation of low-density lipoprotein receptor (LDL-R) in the form of PCSK9/LDL-R complex in lysosomes, which decreases the removal LDL-C from plasma. Evolocumab and Alirocumab, monoclonal antibodies against PCSK9, can effectively reduce the level of LDL-C in patients with statin intolerance or familial hypercholesterolemia, while causing fewer adverse effects and possessing a higher safety profile [6–9]. On the other hand, traditional drugs are limited by their requirement of target protein complex spatial conformation recognition [10]. In addition, conventional drugs cannot target many pathogenic proteins, also known as undruggable proteins, which poses a big challenge to the drug development. Besides, the toxic side effects of statins, especially SAMS, lead to medication non-compliance [6–8]. Moreover, monoclonal antibody drugs (Evolocumab and Alirocumab), targeting hyperlipidemia, need to be administered every two weeks or once a month, whereas statins are taken more frequently. A previous study showed that large doses were required to enhance antibody effectiveness, however, this may produce excessive immune complexes that damage tissues and organs [11]. Finally, due to the expense of monoclonal antibody drugs, long-term and frequent use will bring huge economic burdens to patients and their families [12]. Importantly, traditional small molecule drugs and monoclonal antibodies still cannot reduce the cholesterol to an optimal level in certain patients with familial hypercholesterolemia.
In view of the above points, nucleic acid therapy, especially on an RNA level, may provide more possibilities for the treatment of hyperlipidemia (Figure 1). In terms of drug design, the majority of RNA drugs alter the expression of target gene through Watson-Crick base pairing; unlike traditional drugs, they do not rely on the recognition of the complex spatial conformation of the target proteins [13]. As for patient compliance, RNA drugs have longer half-lives due to special delivery platforms and fewer side effects due to chemical modification. In addition, unlike nucleic acid therapy on the DNA level (genome-editing by CRISPR-Cas9), RNA drugs do not change genetic structure, which results in higher safety in general.

Figure 1. RNA-related therapies mainly include mRNA, ASO (Antisense oligonucleotides), siRNA, microRNA drugs. mRNA drugs can achieve alternative treatment or elicit immune response through synthesizing the corresponding protein. ASOs, composed of 12–25 nucleotides, bind to target-coding RNA or non-coding RNA through the principle of complementary base pairing. This leads to the occupation of translation-related binding sites, which prevents translation without RNA degradation① or the degradation of RNA with ribonuclease H1②. siRNAs are doubled-stranded RNA with guide strands that can form into RISC complexes; they bind to their complementary mRNA and induce target mRNA cleavage, thereby silencing target genes. microRNAs belong to endogenous non-coding RNAs, with 19–25 nucleotides, that inhibit protein translation or promote the degradation of mRNA. microRNA sponges contain identical binding sites to their target microRNAs, which results in decreased binding between the microRNA and 3’-UTR of its target mRNA. Tough Decoy (Tud) is a short hairpin RNA with a stabilized stem-loop and 2 microRNA binding sites; it can greatly reduce target microRNAs (By Figdraw).
2. RNA-based Therapeutic Strategies
2.1. mRNA
It is well known that gene expression is composed of transcription, RNA splicing, translation and post-translational modification. Each step plays a key role in the production of target proteins and have caused a wave of research regarding mRNA-based therapeutics in cancer and vaccine design. The mRNA drugs can affect the production of any protein or peptide in transfected cells, without entering the nucleus to alter DNA sequence.
However, there are three main challenges with mRNA-based therapies, including short half-life, immunogenicity and delivery [10]. In order to minimize the degradation of mRNA by extracellular RNases, 5′-cap, poly (A) tail, 5′- untranslated region (UTR), 3′-UTR and coding regions of mRNA are chemically modified, using cap mimics, poly (A) tail decoration and introduction of stabilizing elements. Immunogenicity is caused by exogenous RNA, with U-rich sequences, which activate Toll-like receptors (e.g. 5-methylcytidine and pseudouridine) [10] and trigger an innate immune response. Another major issue is the delivery of mRNA; there is difficulty crossing the cell membrane due to the large size, negative charge of mRNA and endosome escape. To address the issue, a number of delivery systems with low toxicity have been designed, including polymetric nanoparticles and extracellular vehicles (EVs) [14].
2.2. Antisense Oligonucleotide
Antisense oligonucleotides, composed of 12–25 nucleotides, bind to target-coding RNA (mRNA) or non-coding RNA (e.g., long non-coding RNA and microRNA) through the principle of complementary base pairing; this leads to degradation of RNA under the action of ribonuclease H1 or occupation of the complementary sequences of mRNA, which prevents translation by steric blocking, without RNA degradation [15].
Similar to mRNA-based therapeutics, unmodified ASOs are usually degraded by nucleases in the plasma and cytoplasm, thus they cannot achieve high efficiency. After continuous development, certain modifications have been suggested, which could improve pharmacokinetics and pharmacodynamics, as well as reduce renal toxicity. Cholesterol conjugation can help ASOs interact with serum proteins and lipoproteins, which could promote their entrance into hepatocytes. GalNAc has a high affinity for asialoglyco protein receptor (ASGPR), which is an endocytosed receptor that is predominantly expressed on hepatocytes; ASGPR specifically recognizes and binds with GalNAc. Therefore, cholesterol-GalNAc dual conjugation can be used for delivery of ASOs to the liver to achieve greater target affinity and cellular tropism [16,17].
2.3. Small Interfering RNA
Small interfering RNAs (siRNAs) are doubled-stranded RNAs, which are 20–25 base pairs in length and function in the cytoplasm. Their guide strands incorporate into RNA-induced silencing complex (RISC), bind to the complementary mRNA and induce target mRNA cleavage, thereby silencing target genes. Unlike ASOs, siRNAs can only lead to the degradation of target mRNA, which controls the post-transcriptional event without preventing transcription initiation.
Because of their negative charge, the high sensitivity of nucleases, and the non-specific immunotoxicities induced by siRNA, it is necessary to design a suitable delivery system, which is analogous to mRNA-based therapeutics. Currently, nanoparticle systems have been widely used for drug development, especially lipid nanoparticles (LNPs) [18]. Similar to ASOs, GalNAc conjugation is also used to target hepatocytes in siRNA delivery. However, this conjugation is not effective for mRNA delivery, due to mRNA size is much bigger [19]. In addition, Huang et al. designed a novel siRNA delivery system, the ionizable lipid assisted nucleic acid delivery system (iLAND); this system has been shown to effectively deliver related siRNA to decrease serum cholesterol and triglyceride in high-fat diet (HFD) mice, db/db mice, and human APOC3 transgenic mice [20].
2.4. microRNA
MicroRNAs are conserved non-coding RNAs with 19–25 nucleotides, which participate in various biological events. They are assembled by Ago2 in the RISC complex and bind with the 3’ UTR of mRNA to inhibit its translation or promote mRNA degradation [21]. It has been reported that the pathogenesis of various diseases is attributable to abnormal expression of microRNAs; miR-30c, miR-378a-3p and miR-24 are all involved in pathological mechanisms underlying hyperlipidemia [22–24]. Therefore, developing microRNA inhibitors such as ASO, microRNA sponge and tough decoy (Tud) RNA has become an emerging hotspot in nucleic acid-based therapeutics for hyperlipidemia over the recent years.
microRNA sponges possess binding sites identical to their target microRNA, which result in decreased binding between microRNA and the 3’UTR of its target mRNA. In another word, the more identical binding sites on a microRNA sponge, the higher its inhibition potential [25]. MicroRNA sponges can be integrated into virus vectors, and delivered to targeted organs and cells, depending on the virus serotype and specific promoter. Similar to mRNA delivery, other delivery systems for microRNA sponges, such as LNPs, are also being explored.
Tough Decoy (Tud) is a short hairpin RNA with a stabilized stem-loop and two microRNA binding sites. It has been shown to have great effect in reducing target microRNA; Li et al. successfully reduced miR-320 through miR-320-Tud in db/db and HFD-induced diabetic models [26]. In addition, Dongtak Jeong et al. reported decreased miR-25 level using miR-25-Tud in a mouse model with pressure-overload heart failure; increased SERCA2a protein expression and improved the cardiac function were observed [27]. The delivery strategy of Tud is similar to that of microRNA-sponges, as mentioned above.
3. RNA Therapeutics in Hyperlipidemia
3.1. Antisense oligonucleotides (ASO)
Previous studies showed that increased levels of apolipoprotein C-III in plasma is involved in the process of hypertriglyceridemia [28]. Volanesorsen is an ASO drug targeting ApoC III mRNA to reduce the translation of ApoC-III. The APPROACH trial, a phase III trial in patients with familial chylomicronemia, showed that the triglyceride levels in 77% of participants treated with Volanesorsen were less than 750 mg per deciliter [29]. Although Volanesorsen is very effective in reducing triglyceride level, it leads to an increased risk of thrombocytopenia and other side effects, including injection-site reaction, which impedes the drug approval application. Recently, researchers found that the modification of N-acetylgalactosamine could decrease the toxicity of Volanesoren. Therefore, Olezarsen, a GalNac-conjugated ASO, which targets APOCIII mRNA, has been developed, and no thrombocytopenia has been observed yet [30].
IONIS ANPTL3-LRx is a second-generation ligand-conjugated ASO drug used to target Angptl3 through inhibiting the expression of lipoprotein lipase and endothelial lipase. In the phase I trial of ANGPTL3-LRx, participants receiving ANGPTL3-LRx were divided into two groups: single-dose group and multiple-dose group. Results showed that the levels of Angptl3 protein, VLDL cholesterol, total cholesterol and triglycerides in the multiple-dose group were much lower than the control group. However, in the single-dose group, the aforementioned parameters did not show statistical difference compared to the control group [31].
AZD8233 is an ASO-targeting PCSK9 that can decrease the circulating level of LDL-C. Clinical trials, 2020-000767-23 (EudraCT number) and NCT04964557 (SOLANO), are in phase IIb. In the 2020-000767-23 trial, different dose groups have demonstrated a reduction in the levels of LDL-C and PCSK9 at 12 weeks after subcutaneous injection; injections were administered on Day 1, 8, 29 and 57. NCT04964557 was completed on July 14th, 2022, but the results have not been updated yet.
CiVi 007 is an ASO drug, targeting PCSK9, in phase IIa (NCT04164888). In the trial, participants, with a background of stable statin therapy, were divided into four groups; groups were either receiving placebo or different doses of CiVi 007. CiVi 008 is a third generation PCSK9 antisense molecule for treatment of hypercholesterolemia and atherosclerotic cardiovascular disease. The outcomes of CiVi 007 and CiVi 008 have not been disclosed yet.
3.2. siRNA
PCSK9, which is mainly produced by the liver, mediates the degradation of LDL-R, as mentioned before. Higher level of PCSK9 leads to the onset of hyperlipidemia [32]. Inclisiran is a small interfering RNA; it targets PCSK9 mRNA in hepatocytes to increase the level of LDL-R and promote the degradation of cholesterol. Three clinical trials of Inclisiran, ORION-9 (NCT03397121), ORION-10 (NCT03399370) and ORION-11 (NCT03400800), have been completed. The inclusion criteria for participants were different; the patients in ORION-9 suffered from heterozygous familial hypercholesterolemia, while ORION-10 and -11 enrolled patients with atherosclerotic cardiovascular disease (ASCVD), elevated LDL-C, and were receiving maximally tolerated doses of stains. Inclisiran 300 mg via subcutaneous injection was administered on Day 1 and Day 90, every 6 months, while the same dose of saline solution was administered to the control group. Results from ORION-9, ORION-10 and ORION-11 showed a LDL-C reduction of 49.52%, 57.64% and 53.3% by day 510, respectively. Moreover, the ongoing trial, ORION-16 (NCT04652726), is recruiting participants with heterozygous familial hypercholesterolemia and elevated LDL-C; this trial will be completed in 2025.
4. Perspectives
Hyperlipidemia, especially hypercholesterolemia, has become a worldwide health burden [33]. Although there are many established drugs available for hyperlipidemia, such as statins and monoclonal antibody drugs, each of them has their own limitations, namely intolerable side effects and high costs. Therefore, it is necessary to explore novel treatments for hyperlipidemia. As a result, RNA-based therapeutics has attracted much attention (Table 1).
Table 1 RNA therapeutics in hyperlipidemia.
The GalNAc-conjugated systems used in ASOs and siRNA, as well as LNP delivery, mainly target the liver. The liver is the central hub of lipid metabolism, which suggests great promise for the treatment of hyperlipidemia and other diseases, that originate from the liver, using RNA-based therapeutics. Moreover, various delivery systems and specific chemical modifications, which target different organs and tissues, are needed to obtain better curative effects.
It is worth mentioning that there are some limitations, which need to be addressed, including the efficiency of cellular uptake, endosome escape, off-target effects, and the quality control of modifications and delivery systems. Special delivery systems may be able to solve the issue of drug utilization efficiency, while also decreasing both hepatotoxicity and immunogenicity. A previous study revealed that the incomplete complementary binding between siRNA and mRNA induced off-target effects and hepatoxicity [34]. In addition, quality control is critical in the commercial production of clinical drugs. In a word, it requires continuous efforts from researchers around the world to improve RNA-based therapeutics so that they can play a greater role in the clinical management of hypercholesterolemia.
Author Contributions: Y.Z. conception and drafted this review, C.C. supervised and drafted this review.
Funding: This work was supported by grants from the National Natural Science Foundation of China (82270363 to C.C.). No funding bodies had any role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Acknowledgments: We thank our colleagues in Dr. Chen's group for technical assistance and stimulating discussions during the course of this investigation.
Conflicts of Interest: The authors declare there is no conflict of interests.
Introduction
Hyperlipidemia has become a common health burden over recent decades, as patients have increasing levels of cholesterol, triglycerides and/or lipoproteins. It is recognized as a risk factor for type 2 diabetes, pancreatitis, non-alcoholic fatty liver disease, chronic kidney disease and cardiovascular diseases [1]. Previous studies showed that lipid metabolism consists of de novo lipogenesis, as well as uptake, oxidation and degradation of lipids, such as fatty acid, triglycerides and cholesterol. Their transportation and degradation are greatly affected by the lipoprotein metabolism, including low-density lipoprotein (LDL), very-low-density lipoprotein (VLDL) and high-density lipoprotein (HDL).
The traditional drugs against hyperlipidemia are mainly small molecule drugs, especially statins and monoclonal antibody drugs. Statins are the first-line lipid-lowering drugs in clinical practice nowadays; they lower the level of LDL-C by inhibiting 3-hydroxy-3-methyl-glutaryl coenzyme-A reductase, a rate-limiting enzyme in cholesterol synthesis. However, with long-term use, some patients suffer from adverse effects; the main reasons for statin intolerance are liver injury [2], statin-associated muscle symptoms (SAMS) [3] and new-onset type 2 diabetes (NOD) [4,5]. Previous studies identified that proprotein convertase subtilisin/kexin type 9 (PCSK9), a serine protease synthesized primarily in hepatocytes, was of great importance to the pathogenesis of familial hypercholesterolemia. PCSK9 acts by inducing the degradation of low-density lipoprotein receptor (LDL-R) in the form of PCSK9/LDL-R complex in lysosomes, which decreases the removal LDL-C from plasma. Evolocumab and Alirocumab, monoclonal antibodies against PCSK9, can effectively reduce the level of LDL-C in patients with statin intolerance or familial hypercholesterolemia, while causing fewer adverse effects and possessing a higher safety profile [6–9]. On the other hand, traditional drugs are limited by their requirement of target protein complex spatial conformation recognition [10]. In addition, conventional drugs cannot target many pathogenic proteins, also known as undruggable proteins, which poses a big challenge to the drug development. Besides, the toxic side effects of statins, especially SAMS, lead to medication non-compliance [6–8]. Moreover, monoclonal antibody drugs (Evolocumab and Alirocumab), targeting hyperlipidemia, need to be administered every two weeks or once a month, whereas statins are taken more frequently. A previous study showed that large doses were required to enhance antibody effectiveness, however, this may produce excessive immune complexes that damage tissues and organs [11]. Finally, due to the expense of monoclonal antibody drugs, long-term and frequent use will bring huge economic burdens to patients and their families [12]. Importantly, traditional small molecule drugs and monoclonal antibodies still cannot reduce the cholesterol to an optimal level in certain patients with familial hypercholesterolemia.
In view of the above points, nucleic acid therapy, especially on an RNA level, may provide more possibilities for the treatment of hyperlipidemia (Figure 1). In terms of drug design, the majority of RNA drugs alter the expression of target gene through Watson-Crick base pairing; unlike traditional drugs, they do not rely on the recognition of the complex spatial conformation of the target proteins [13]. As for patient compliance, RNA drugs have longer half-lives due to special delivery platforms and fewer side effects due to chemical modification. In addition, unlike nucleic acid therapy on the DNA level (genome-editing by CRISPR-Cas9), RNA drugs do not change genetic structure, which results in higher safety in general.

Figure 1. RNA-related therapies mainly include mRNA, ASO (Antisense oligonucleotides), siRNA, microRNA drugs. mRNA drugs can achieve alternative treatment or elicit immune response through synthesizing the corresponding protein. ASOs, composed of 12–25 nucleotides, bind to target-coding RNA or non-coding RNA through the principle of complementary base pairing. This leads to the occupation of translation-related binding sites, which prevents translation without RNA degradation① or the degradation of RNA with ribonuclease H1②. siRNAs are doubled-stranded RNA with guide strands that can form into RISC complexes; they bind to their complementary mRNA and induce target mRNA cleavage, thereby silencing target genes. microRNAs belong to endogenous non-coding RNAs, with 19–25 nucleotides, that inhibit protein translation or promote the degradation of mRNA. microRNA sponges contain identical binding sites to their target microRNAs, which results in decreased binding between the microRNA and 3’-UTR of its target mRNA. Tough Decoy (Tud) is a short hairpin RNA with a stabilized stem-loop and 2 microRNA binding sites; it can greatly reduce target microRNAs (By Figdraw).
RNA-based Therapeutic Strategies
mRNA
It is well known that gene expression is composed of transcription, RNA splicing, translation and post-translational modification. Each step plays a key role in the production of target proteins and have caused a wave of research regarding mRNA-based therapeutics in cancer and vaccine design. The mRNA drugs can affect the production of any protein or peptide in transfected cells, without entering the nucleus to alter DNA sequence.
However, there are three main challenges with mRNA-based therapies, including short half-life, immunogenicity and delivery [10]. In order to minimize the degradation of mRNA by extracellular RNases, 5′-cap, poly (A) tail, 5′- untranslated region (UTR), 3′-UTR and coding regions of mRNA are chemically modified, using cap mimics, poly (A) tail decoration and introduction of stabilizing elements. Immunogenicity is caused by exogenous RNA, with U-rich sequences, which activate Toll-like receptors (e.g. 5-methylcytidine and pseudouridine) [10] and trigger an innate immune response. Another major issue is the delivery of mRNA; there is difficulty crossing the cell membrane due to the large size, negative charge of mRNA and endosome escape. To address the issue, a number of delivery systems with low toxicity have been designed, including polymetric nanoparticles and extracellular vehicles (EVs) [14].
Antisense Oligonucleotide
Antisense oligonucleotides, composed of 12–25 nucleotides, bind to target-coding RNA (mRNA) or non-coding RNA (e.g., long non-coding RNA and microRNA) through the principle of complementary base pairing; this leads to degradation of RNA under the action of ribonuclease H1 or occupation of the complementary sequences of mRNA, which prevents translation by steric blocking, without RNA degradation [15].
Similar to mRNA-based therapeutics, unmodified ASOs are usually degraded by nucleases in the plasma and cytoplasm, thus they cannot achieve high efficiency. After continuous development, certain modifications have been suggested, which could improve pharmacokinetics and pharmacodynamics, as well as reduce renal toxicity. Cholesterol conjugation can help ASOs interact with serum proteins and lipoproteins, which could promote their entrance into hepatocytes. GalNAc has a high affinity for asialoglyco protein receptor (ASGPR), which is an endocytosed receptor that is predominantly expressed on hepatocytes; ASGPR specifically recognizes and binds with GalNAc. Therefore, cholesterol-GalNAc dual conjugation can be used for delivery of ASOs to the liver to achieve greater target affinity and cellular tropism [16,17].
Small Interfering RNA
Small interfering RNAs (siRNAs) are doubled-stranded RNAs, which are 20–25 base pairs in length and function in the cytoplasm. Their guide strands incorporate into RNA-induced silencing complex (RISC), bind to the complementary mRNA and induce target mRNA cleavage, thereby silencing target genes. Unlike ASOs, siRNAs can only lead to the degradation of target mRNA, which controls the post-transcriptional event without preventing transcription initiation.
Because of their negative charge, the high sensitivity of nucleases, and the non-specific immunotoxicities induced by siRNA, it is necessary to design a suitable delivery system, which is analogous to mRNA-based therapeutics. Currently, nanoparticle systems have been widely used for drug development, especially lipid nanoparticles (LNPs) [18]. Similar to ASOs, GalNAc conjugation is also used to target hepatocytes in siRNA delivery. However, this conjugation is not effective for mRNA delivery, due to mRNA size is much bigger [19]. In addition, Huang et al. designed a novel siRNA delivery system, the ionizable lipid assisted nucleic acid delivery system (iLAND); this system has been shown to effectively deliver related siRNA to decrease serum cholesterol and triglyceride in high-fat diet (HFD) mice, db/db mice, and human APOC3 transgenic mice [20].
microRNA
MicroRNAs are conserved non-coding RNAs with 19–25 nucleotides, which participate in various biological events. They are assembled by Ago2 in the RISC complex and bind with the 3’ UTR of mRNA to inhibit its translation or promote mRNA degradation [21]. It has been reported that the pathogenesis of various diseases is attributable to abnormal expression of microRNAs; miR-30c, miR-378a-3p and miR-24 are all involved in pathological mechanisms underlying hyperlipidemia [22–24]. Therefore, developing microRNA inhibitors such as ASO, microRNA sponge and tough decoy (Tud) RNA has become an emerging hotspot in nucleic acid-based therapeutics for hyperlipidemia over the recent years.
microRNA sponges possess binding sites identical to their target microRNA, which result in decreased binding between microRNA and the 3’UTR of its target mRNA. In another word, the more identical binding sites on a microRNA sponge, the higher its inhibition potential [25]. MicroRNA sponges can be integrated into virus vectors, and delivered to targeted organs and cells, depending on the virus serotype and specific promoter. Similar to mRNA delivery, other delivery systems for microRNA sponges, such as LNPs, are also being explored.
Tough Decoy (Tud) is a short hairpin RNA with a stabilized stem-loop and two microRNA binding sites. It has been shown to have great effect in reducing target microRNA; Li et al. successfully reduced miR-320 through miR-320-Tud in db/db and HFD-induced diabetic models [26]. In addition, Dongtak Jeong et al. reported decreased miR-25 level using miR-25-Tud in a mouse model with pressure-overload heart failure; increased SERCA2a protein expression and improved the cardiac function were observed [27]. The delivery strategy of Tud is similar to that of microRNA-sponges, as mentioned above.
RNA Therapeutics in Hyperlipidemia
Antisense oligonucleotides (ASO)
Volanesorsen and Olezarsen
Previous studies showed that increased levels of apolipoprotein C-III in plasma is involved in the process of hypertriglyceridemia [28]. Volanesorsen is an ASO drug targeting ApoC III mRNA to reduce the translation of ApoC-III. The APPROACH trial, a phase III trial in patients with familial chylomicronemia, showed that the triglyceride levels in 77% of participants treated with Volanesorsen were less than 750 mg per deciliter [29]. Although Volanesorsen is very effective in reducing triglyceride level, it leads to an increased risk of thrombocytopenia and other side effects, including injection-site reaction, which impedes the drug approval application. Recently, researchers found that the modification of N-acetylgalactosamine could decrease the toxicity of Volanesoren. Therefore, Olezarsen, a GalNac-conjugated ASO, which targets APOCIII mRNA, has been developed, and no thrombocytopenia has been observed yet [30].
IONIS ANPTL3-LRx (ISIS 703802)
IONIS ANPTL3-LRx is a second-generation ligand-conjugated ASO drug used to target Angptl3 through inhibiting the expression of lipoprotein lipase and endothelial lipase. In the phase I trial of ANGPTL3-LRx, participants receiving ANGPTL3-LRx were divided into two groups: single-dose group and multiple-dose group. Results showed that the levels of Angptl3 protein, VLDL cholesterol, total cholesterol and triglycerides in the multiple-dose group were much lower than the control group. However, in the single-dose group, the aforementioned parameters did not show statistical difference compared to the control group [31].
AZD8233
AZD8233 is an ASO-targeting PCSK9 that can decrease the circulating level of LDL-C. Clinical trials, 2020-000767-23 (EudraCT number) and NCT04964557 (SOLANO), are in phase IIb. In the 2020-000767-23 trial, different dose groups have demonstrated a reduction in the levels of LDL-C and PCSK9 at 12 weeks after subcutaneous injection; injections were administered on Day 1, 8, 29 and 57. NCT04964557 was completed on July 14th, 2022, but the results have not been updated yet.
CiVi 007 and CiVi 008
CiVi 007 is an ASO drug, targeting PCSK9, in phase IIa (NCT04164888). In the trial, participants, with a background of stable statin therapy, were divided into four groups; groups were either receiving placebo or different doses of CiVi 007. CiVi 008 is a third generation PCSK9 antisense molecule for treatment of hypercholesterolemia and atherosclerotic cardiovascular disease. The outcomes of CiVi 007 and CiVi 008 have not been disclosed yet.
siRNA
ALN-PCSsc (Inclisiran)
PCSK9, which is mainly produced by the liver, mediates the degradation of LDL-R, as mentioned before. Higher level of PCSK9 leads to the onset of hyperlipidemia [32]. Inclisiran is a small interfering RNA; it targets PCSK9 mRNA in hepatocytes to increase the level of LDL-R and promote the degradation of cholesterol. Three clinical trials of Inclisiran, ORION-9 (NCT03397121), ORION-10 (NCT03399370) and ORION-11 (NCT03400800), have been completed. The inclusion criteria for participants were different; the patients in ORION-9 suffered from heterozygous familial hypercholesterolemia, while ORION-10 and -11 enrolled patients with atherosclerotic cardiovascular disease (ASCVD), elevated LDL-C, and were receiving maximally tolerated doses of stains. Inclisiran 300 mg via subcutaneous injection was administered on Day 1 and Day 90, every 6 months, while the same dose of saline solution was administered to the control group. Results from ORION-9, ORION-10 and ORION-11 showed a LDL-C reduction of 49.52%, 57.64% and 53.3% by day 510, respectively. Moreover, the ongoing trial, ORION-16 (NCT04652726), is recruiting participants with heterozygous familial hypercholesterolemia and elevated LDL-C; this trial will be completed in 2025.
Perspectives
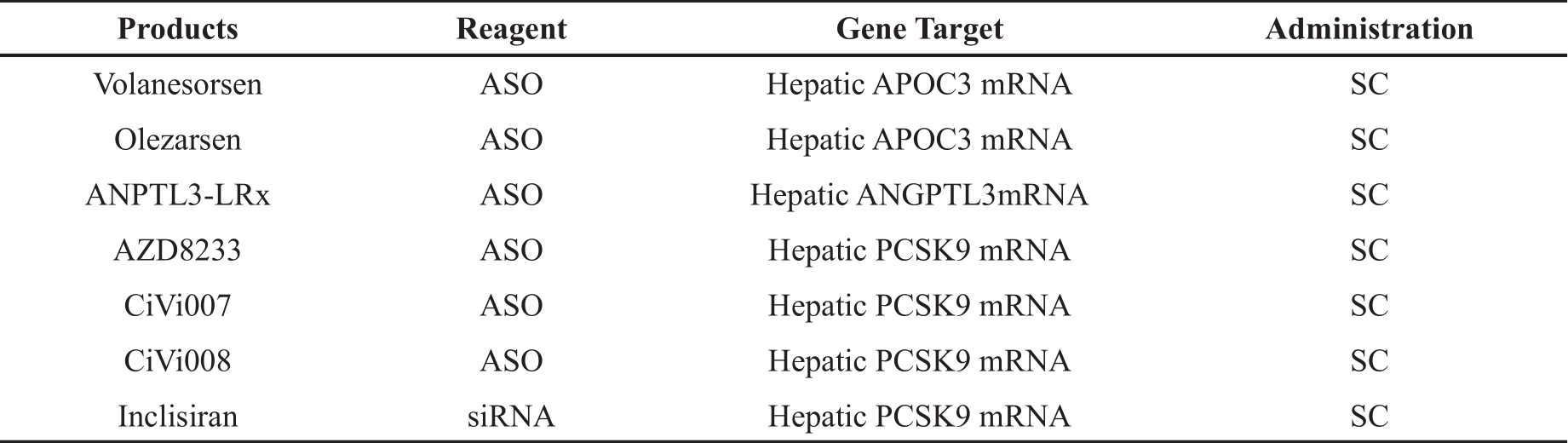
Hyperlipidemia, especially hypercholesterolemia, has become a worldwide health burden [33]. Although there are many established drugs available for hyperlipidemia, such as statins and monoclonal antibody drugs, each of them has their own limitations, namely intolerable side effects and high costs. Therefore, it is necessary to explore novel treatments for hyperlipidemia. As a result, RNA-based therapeutics has attracted much attention (Table 1).
| Products | Reagent | Gene Target | Administration |
|---|---|---|---|
| Volanesorsen | ASO | Hepatic APOC3 mRNA | SC |
| Olezarsen | ASO | Hepatic APOC3 mRNA | SC |
| ANPTL3-LRx | ASO | Hepatic ANGPTL3mRNA | SC |
| AZD8233 | ASO | Hepatic PCSK9 mRNA | SC |
| CiVi007 | ASO | Hepatic PCSK9 mRNA | SC |
| CiVi008 | ASO | Hepatic PCSK9 mRNA | SC |
| Inclisiran | siRNA | Hepatic PCSK9 mRNA | SC |
The GalNAc-conjugated systems used in ASOs and siRNA, as well as LNP delivery, mainly target the liver. The liver is the central hub of lipid metabolism, which suggests great promise for the treatment of hyperlipidemia and other diseases, that originate from the liver, using RNA-based therapeutics. Moreover, various delivery systems and specific chemical modifications, which target different organs and tissues, are needed to obtain better curative effects.
It is worth mentioning that there are some limitations, which need to be addressed, including the efficiency of cellular uptake, endosome escape, off-target effects, and the quality control of modifications and delivery systems. Special delivery systems may be able to solve the issue of drug utilization efficiency, while also decreasing both hepatotoxicity and immunogenicity. A previous study revealed that the incomplete complementary binding between siRNA and mRNA induced off-target effects and hepatoxicity [34]. In addition, quality control is critical in the commercial production of clinical drugs. In a word, it requires continuous efforts from researchers around the world to improve RNA-based therapeutics so that they can play a greater role in the clinical management of hypercholesterolemia.
References
- Rosenson R.S.; Najera S.D.; Hegele R.A. Heterozygous familial hypercholesterolemia presenting as chylomicronemia syndrome. J. Clin. Lipidol., 2017, 11(1): 294-296. DOI: https://doi.org/10.1016/j.jacl.2016.12.005
- Averbukh L.D.; Turshudzhyan A.; Wu D.C.; et al. Statin-induced liver injury patterns: a clinical review. J Clin Transl Hepatol, 2022, 10(3): 543-552. DOI: https://doi.org/10.14218/JCTH.2021.00271
- Vinci P.; Panizon E.; Tosoni L.M.; et al. Statin-associated myopathy: emphasis on mechanisms and targeted therapy. Int. J. Mol. Sci., 2021, 22(21): 11687. DOI: https://doi.org/10.3390/ijms222111687
- Carmena R.; Betteridge D.J. Diabetogenic action of statins: mechanisms. Curr. Atheroscler. Rep., 2019, 21(6): 23. DOI: https://doi.org/10.1007/s11883-019-0780-z
- Galicia-Garcia U.; Jebari S.; Larrea-Sebal A.; et al. Statin treatment-induced development of type 2 diabetes: from clinical evidence to mechanistic insights. Int. J. Mol. Sci., 2020, 21(13): 4725. DOI: https://doi.org/10.3390/ijms21134725
- Santos R.D.; Stein E.A.; Hovingh G.K.; et al. Long-term evolocumab in patients with familial hypercholesterolemia. J. Am. Coll. Cardiol., 2020, 75(6): 565-574. DOI: https://doi.org/10.1016/j.jacc.2019.12.020
- Santos R.D.; Ruzza A.; Hovingh G.K.; et al. Evolocumab in pediatric heterozygous familial hypercholesterolemia. N. Engl. J. Med., 2020, 383(14): 1317-1327. DOI: https://doi.org/10.1056/NEJMoa2019910
- Blom D.J.; Harada-Shiba M.; Rubba P.; et al. Efficacy and safety of alirocumab in adults with homozygous familial hypercholesterolemia: the ODYSSEY HoFH trial. J. Am. Coll. Cardiol., 2020, 76(2): 131-142. DOI: https://doi.org/10.1016/j.jacc.2020.05.027
- Nissen S.E.; Stroes E.; Dent-Acosta R.E.; et al. Efficacy and tolerability of evolocumab vs ezetimibe in patients with muscle-related statin intolerance: the GAUSS-3 randomized clinical trial. JAMA, 2016, 315(15): 1580-1590. DOI: https://doi.org/10.1001/jama.2016.3608
- Hu B.; Zhong L.P.; Weng Y.H.; et al. Therapeutic siRNA: state of the art. Signal Transduction Targeted Ther., 2020, 5(1): 101. DOI: https://doi.org/10.1038/s41392-020-0207-x
- Tsimikas S. RNA-targeted therapeutics for lipid disorders. Curr. Opin. Lipidol., 2018, 29(6): 459-466. DOI: https://doi.org/10.1097/MOL.0000000000000549
- Kazi D.S.; Penko J.; Coxson P.G.; et al. Updated cost-effectiveness analysis of PCSK9 inhibitors based on the results of the FOURIER trial. JAMA, 2017, 318(8): 748-750. DOI: https://doi.org/10.1001/jama.2017.9924
- Yu A.M.; Choi Y.H.; Tu M.J. RNA drugs and RNA targets for small molecules: principles, progress, and challenges. Pharmacol. Rev., 2020, 72(4): 862-898. DOI: https://doi.org/10.1124/pr.120.019554
- Qin S.G.; Tang X.S.; Chen Y.T.; et al. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduction Targeted Ther., 2022, 7(1): 166. DOI: https://doi.org/10.1038/s41392-022-01007-w
- Katzmann J.L.; Packard C.J.; Chapman M.J.; et al. Targeting RNA with antisense oligonucleotides and small interfering RNA: JACC state-of-the-art review. J. Am. Coll. Cardiol., 2020, 76(5): 563-579. DOI: https://doi.org/10.1016/j.jacc.2020.05.070
- Kulkarni J.A.; Witzigmann D.; Thomson S.B.; et al. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol., 2021, 16(6): 630-643. DOI: https://doi.org/10.1038/s41565-021-00898-0
- Wada F.; Yamamoto T.; Ueda T.; et al. Cholesterol-GalNAc dual conjugation strategy for reducing renal distribution of antisense oligonucleotides. Nucleic Acid Ther., 2018, 28(1): 50-57. DOI: https://doi.org/10.1089/nat.2017.0698
- Kubczak M.; Michlewska S.; Bryszewska M.; et al. Nanoparticles for local delivery of siRNA in lung therapy. Adv. Drug Delivery Rev., 2021, 179: 114038. DOI: https://doi.org/10.1016/j.addr.2021.114038
- Weng Y.H.; Li C.H.; Yang T.R.; et al. The challenge and prospect of mRNA therapeutics landscape. Biotechnol. Adv., 2020, 40: 107534. DOI: https://doi.org/10.1016/j.biotechadv.2020.107534
- Hu B.; Li B.; Li K.; et al. Thermostable ionizable lipid-like nanoparticle (iLAND) for RNAi treatment of hyperlipidemia. Sci. Adv., 2022, 8(7): eabm1418. DOI: https://doi.org/10.1126/sciadv.abm1418
- Ammirati E.; Veronese G.; Brambatti M.; et al. Fulminant versus acute nonfulminant myocarditis in patients with left ventricular systolic dysfunction. J. Am. Coll. Cardiol., 2019, 74(3): 299-311.
- Soh J.; Iqbal J.; Queiroz J.; et al. MicroRNA-30c reduces hyperlipidemia and atherosclerosis in mice by decreasing lipid synthesis and lipoprotein secretion. Nat. Med., 2013, 19(7): 892-900. DOI: https://doi.org/10.1038/nm.3200
- Zhang T.P.; Shi H.T.; Liu N.N.; et al. Activation of microRNA-378a-3p biogenesis promotes hepatic secretion of VLDL and hyperlipidemia by modulating ApoB100-Sortilin1 axis. Theranostics, 2020, 10(9): 3952-3966. DOI: https://doi.org/10.7150/thno.39578
- Ng R.; Wu H.; Xiao H.; et al. Inhibition of microRNA-24 expression in liver prevents hepatic lipid accumulation and hyperlipidemia. Hepatology, 2014, 60(2): 554-564. DOI: https://doi.org/10.1002/hep.27153
- Bernardo B.C.; Gregorevic P.; Ritchie R.H.; et al. Generation of MicroRNA-34 sponges and tough decoys for the heart: developments and challenges. Front. Pharmacol., 2018, 9: 1090. DOI: https://doi.org/10.3389/fphar.2018.01090
- Li H.P.; Fan J.H.; Zhao Y.R.; et al. Nuclear miR-320 mediates diabetes-induced cardiac dysfunction by activating transcription of fatty acid metabolic genes to cause lipotoxicity in the heart. Circ. Res., 2019, 125(12): 1106-1120. DOI: https://doi.org/10.1161/CIRCRESAHA.119.314898
- Jeong D.; Yoo J.; Lee P.; et al. miR-25 tough decoy enhances cardiac function in heart failure. Mol. Ther., 2018, 26(3): 718-729. DOI: https://doi.org/10.1016/j.ymthe.2017.11.014
- Zheng C.Y.; Khoo C.; Furtado J.; et al. Apolipoprotein C-Ⅲ and the metabolic basis for hypertriglyceridemia and the dense low-density lipoprotein phenotype. Circulation, 2010, 121(15): 1722-1734. DOI: https://doi.org/10.1161/CIRCULATIONAHA.109.875807
- Witztum J.L.; Gaudet D.; Freedman S.D.; et al. Volanesorsen and triglyceride levels in familial chylomicronemia syndrome. N. Engl. J. Med., 2019, 381(6): 531-542. DOI: https://doi.org/10.1056/NEJMoa1715944
- Hegele R.A. Apolipoprotein C-Ⅲ inhibition to lower triglycerides: one ring to rule them all?. Eur. Heart J., 2022, 43(14): 1413-1415. DOI: https://doi.org/10.1093/eurheartj/ehab890
- Graham M.J.; Lee R.G.; Brandt T.A.; et al. Cardiovascular and metabolic effects of ANGPTL3 antisense oligonucleotides. N. Engl. J. Med., 2017, 377(3): 222-232. DOI: https://doi.org/10.1056/NEJMoa1701329
- Horton J.D.; Cohen J.C.; Hobbs H.H. Molecular biology of PCSK9: its role in LDL metabolism. Trends Biochem. Sci., 2007, 32(2): 71-77. DOI: https://doi.org/10.1016/j.tibs.2006.12.008
- Collaboration NCDRF. Repositioning of the global epicentre of non-optimal cholesterol. Nature, 2020, 582(7810): 73-77. DOI: https://doi.org/10.1038/s41586-020-2338-1
- Blanco-Domínguez R.; Sánchez-Díaz R.; De La Fuente H.; et al. A novel circulating MicroRNA for the detection of acute myocarditis. N. Engl. J. Med., 2021, 384(21): 2014-2027.


